Annotated bibliography 1:
Seibel, B.A., Chausson, F., Lallier, F.H., Zal, F. And Childress, J.J. 1999. Vampire blood: respiratory physiology of the vampire squid (Cephalopoda: Vampyromorpha: Vampyroteuthidae) in relation to the oxygen minimum layer. Experimental Biology Online. 4:1-10.
The study was conducted to examine the physiological adaptations of the Vampire Squid which allow it to survive in an extreme environment (minimal oxygen) where other species of Cephalopods and Invertebrates cannot. Seibel et al. hypothesized that the hemocyanin of Vampire Squid must be extremely high and also that it’s affinity to oxygen must be much higher than that of other cephalopods, to be able to survive.
The Vampire Squid is the only cephalopod able to carry out its entire life cycle in the core of the oxygen minimal zone. Pelagic crustaceans in the OMA have large ventilatory volume, large gill surface areas, short diffusion distances for oxygen to travel into the blood, and hemocyanin proteins with high oxygen affinities. Vampire Squids have moderate gill surface area by comparison as well as moderate diffusion distances and the connection between ventilation and locomotion prevents consistent high respiratory rates. The Vampire Squid must be physiological adapted in such a way as to be able to survive in these zones, and therefore and therefore the hemocyanin oxygen affinities must be important.
Specimens were collected off the coast of California and blood was collected and frozen. Oxygen dissociation curves were established by adjusting the pH of the blood as well as a diffusion chamber was used to regulate oxygen levels, and the ionic components/concentrations of the blood were determined. The hemocyanin-oxygen affinity measured was extremely high, and higher than any other cephalopod.
The extremely low oxygen content of the waters that the Vampire Squid inhabit (OMZs) requires that this species must have efficient and effective absorption of oxygen from the surrounding environment. Vampire Squid’s hemocyanin was found to have the highest oxygen affinity of any cephalopod. The efficiency of this hemocyanin was found to be one of the most important physiological adaptations of Vampire Squid to the OMZs.
Annotated bibliography 2:
Seibel, B.A., Thuesen, E.V. and Childress, J.J. 1998. Flight of the Vampire: Ontogenetic Gait-Transition in Vampyroteuthis infernalis (Cephalopoda: Vampyromorpha: Vampyroteuthidae). The Journal of Experimental Biology. 201:2413-2424.
The goal of this study was to investigate a change in swimming method over the course of the Vampire Squid’s life cycle and its relation to efficiency in the oxygen minimal zone. It was examined whether or not there was a relationship between the size of the organism and the efficiency of fin versus jet-propulsion swimming as well as whether or such relationships can result in transitions in locomotion through ontogeny.
Fin swimming is more common in adult Vampire Squid than jet-propulsion, due to its energetic efficiency. However, juvenile Vampire Squid may rely heavily on jet-propulsion and eventually undergo a metamorphosis which results in a change in fin position, relative size and shape as the individual becomes an adult and transitions to fin swimming.
Citrate synthase (CS) and octopine dehydrogenase (ODH) are useful indicators of aerobic and anaerobic metabolism and can be used to determine where the main areas of muscle contractions (for locomotion) are concentrated. Vampire Squid specimens were captured and the shape, size and position of the fins was recorded. CS and ODH levels were measured in the fin muscles (fin swimming) and mantle muscles (jet-propulsion) in juveniles and adults. High levels were shown in the fin muscle in adults, and high levels were shown in the mantle muscle in juveniles.
Changes from jet-propulsion to fin swimming were associated with a change in fin shape, size and position due to metamorphosis from juvenile to adult. Therefore there is a relationship between size of an organism and most efficient swimming methods. The transition between swimming styles can be defined as an “ontogenetic gait-transition”.
Annotated bibliography 3:
Bonnaud, L., Boucher-Rodoni, R. and Monnerott, M. 1997. Phylogeny of Cephalopods Inferred from Mitochondrial DNA Sequences. Molecular Phylogenetics and Evolution. 7(1):44-54.
The class Cephalopoda, of the Phylum Mollusca, was divided into groups primarily based on morphological and paleontological characters. Many of the relationships within this group, such as the relationship between Vampire Squid (Vampyromorpha) and Decapods/Octopods. The purpose of this study was to determine the accuracy of previous phylogenetic studies.
Cephalopoda is currently divided into Nautiloidea (Nautiloids) and Coleoidea (Decapods – squid and cuttlefish, Octopods – Octopi). The monophyly of many groups, such as the decapods and groups within the decapods, was not sufficiently supported by morphological evidence alone. For example: Vampire Squid have characters similar to both decapods and octopods, and may be a sister group to either. Mitochondrial DNA variation can be used to infer phylogenetic relationships, and was examined in this study.
Specimens of numerous groups of Cephalopods were collected and the mtDNA was isolated from tissue homogenates. PCR amplification was used on the mtDNA and these samples were sequenced. Phylogenetic analyses were carried out on the mtDNA sequences to determine the relationships between the organisms being studied. The monophyly of decapods was confirmed and Vampyromorpha was identified as being a sister group to all other octopods.
Some of the relationships between Cephalopods (as determined by morphological and paleontological data) were determined to be inaccurate and others were more extensively resolved. The Vampire Squid was placed as a sister group to Octopods, and not within the Decapods, despite its confusing morphological characters.






 The bright points on this image are the arm tip and fin base photophores, lit up in response to the presence of a dangerous predator (the camera).
The bright points on this image are the arm tip and fin base photophores, lit up in response to the presence of a dangerous predator (the camera).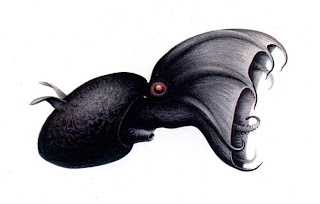 This is the what the Vampire Squid from the original description of this species looked like. It was described as "a very small but terrible octopus, black as night, with ivory white jaws and blood red eyes". As you can see from the following images, this description could not be farther from the truth.
This is the what the Vampire Squid from the original description of this species looked like. It was described as "a very small but terrible octopus, black as night, with ivory white jaws and blood red eyes". As you can see from the following images, this description could not be farther from the truth. The terrible "Vampire Squid from Hell".
The terrible "Vampire Squid from Hell". The massive eye of the Vampire Squid. Like other cephalopod eyes, the Vampire Squid's eyes are complex and highly evolved. The are not totally unlike our own eyes.
The massive eye of the Vampire Squid. Like other cephalopod eyes, the Vampire Squid's eyes are complex and highly evolved. The are not totally unlike our own eyes.